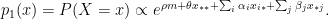In the last group meeting I reported on the progress that I have made regarding the development of a protocol for the systematic use of Natural Move Monte Carlo simulations.
Natural Move Monte Carlo simulations
Natural Moves are degrees of freedom that describe the collective motion of groups of residues. In DNA this might be the concerted motion of a double helix; in proteins this could be the movement of a stable secondary structure element such as a beta-sheet. These segments are joined by so called melting areas. At each simulation step the segments are propagated independently in an MC fashion. The resulting chain breaks are resolved by a chain closure algorithm that acts on the melting areas. This results in a reduction of degrees of freedom of several orders of magnitude. Therefore, large complexes and conformational changes can be sampled more effectively.
In order to get sensible results, however, the initial decomposition of the system is important. The challenge is to accurately represent the plasticity of the system, while keeping the number of degrees of freedom as small as possible. Detailed insight into the flexibility of the system might be gained from experimental sources such as NMR or computational methods such as MD simulations and Normal Mode Analysis. This can help with defining segments and melting areas. However, there are many systems for which this data is not available. Even if it is, there is no guarantee that the segmentation is correct.
Therefore, I am developing a protocol that allows for the evaluation of a range of different test cases that each reflect a unique set of segments and melting areas.
Augmented Modelling Protocol
This protocol is aimed at the systematic evaluation of NMMC segmentations. It allows researchers to feed experimental information, biological knowledge and educated guesses into molecular simulations and so provides a framework for testing competing hypotheses. The protocol has four steps.
Step 1: Segmentation of the system into low-level segments
The initial segmentation contains all possible areas of flexibility that may play a role in conformational changes in the system of interest. This decision may be influenced by many sources. For now, however, we only consider secondary structure information. Helices and beta strands are treated as potential segments. Unstructured regions such as kinks, loops and random coils are treated as melting areas. For a small fold with four helices we get the segmentation shown in figure 1a.
Step 2: Formulate test cases
Generate multiple test cases that reflect hypotheses about the mechanism of interest. In this step we try to narrow down the degrees of freedom as much as possible in order to retain sampling efficiency. This is done by selectively deactivating some melting areas that were defined in step 1. For a system with three melting areas that can either be on or off, 2^3 = 8 different test cases may be generated (example shown in figure 1b).

Figure 1 a) Segmentation of a small α-fold. The blue rectangles represent α-helices. The dashed lines indicate the presence of melting areas I, II and III. Each melting area can be switched on or off (1/0) b) Example of a test case in which the first of three melting area is switched off. c) The six degrees of freedom along which a segment is propagated.
Step 3: Perform simulations
Sample the conformational space of all test cases that were generated in step 2. We generally use Parallel Tempering or Simulated Tempering algorithm to accelerate the sampling process. These methods rely on the modulation of temperature to overcome energy barriers.
Step 4: Evaluate results
Score the results against a given control and rank the test cases accordingly. The scoring might be done by comparing experimental distributions of observables with those generated by simulations (e.g. Kullback-Leibler divergence). A test case that reproduces desired expectation values of observables might then be considered as a candidate hypothesis for a certain structural mechanism.
What’s next?
I am currently working on example uses for this protocol. These include questions regarding aspects of protein folding and the stability of the empty MHC II binding groove.
![Example of a simple, link-clustered network. Nodes can be seen to be in multiple communities as edges are assigned to communities (c.f. node 4 in red, green and yellow communities). [Ahn et al., 2010]](https://i0.wp.com/www.blopig.com/blog/wp-content/uploads/2014/10/link_clustering_graph_partition.png?ssl=1)

 represents the total number of edges in the network and
represents the total number of edges in the network and 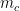 and
and  represent the number of edges and nodes in the community
represent the number of edges and nodes in the community  respectively.
respectively.![Schematic Diagram showing how the similarity between edges e_ik and and e_jk is calculated in Link Clustering. The overlap of the inclusive neighbour sets of nodes i and j is divided by the union of these sets. The inclusive neighbour set of node i is computed by taking the neighbours of node i and including node i itself. The nodes in grey are overlapping between both sets, therefore the similarity between edges e_ik and e_jk is 4/12 or 1/3. [Ahn et al 2010]](https://i0.wp.com/www.blopig.com/blog/wp-content/uploads/2014/10/link_clustering_edge_similarity.png?ssl=1)
 and and
and and 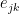 is calculated in Link Clustering. The overlap of the inclusive neighbour sets of nodes i and j is divided by the union of these sets. The inclusive neighbour set of node i is computed by taking the neighbours of node i and including node i itself. The nodes in grey are overlapping between both sets, therefore the similarity between edges
is calculated in Link Clustering. The overlap of the inclusive neighbour sets of nodes i and j is divided by the union of these sets. The inclusive neighbour set of node i is computed by taking the neighbours of node i and including node i itself. The nodes in grey are overlapping between both sets, therefore the similarity between edges ![Schematic of the Affiliation Graph Model, which shows nodes (coloured squares) being affiliated with communities (circles A, B, and C) as shown by the arrows. Nodes can have multiple community affiliations as indicated by red and purple node squares. The probability of interaction between two nodes which are both in only community A, is p_A. [Yang and Leskovec, 2012]](https://i0.wp.com/www.blopig.com/blog/wp-content/uploads/2014/10/AGM_model.png?ssl=1)
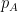 . [Yang and Leskovec, 2012]
. [Yang and Leskovec, 2012]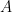 . For nodes
. For nodes  and
and  , which may share several community affiliations, the probability of interaction,
, which may share several community affiliations, the probability of interaction,  is given by the equation:
is given by the equation:
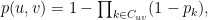
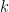 denotes a community in the set of communities shared by nodes
denotes a community in the set of communities shared by nodes  . This model is be fitted to the network using a Metropolis-Hastings algorithm with a Markov chain constructed by pre-defined defined steps in the space of possible Community Affiliation Matrices (c.f. Yang and Leskovec 2012).
. This model is be fitted to the network using a Metropolis-Hastings algorithm with a Markov chain constructed by pre-defined defined steps in the space of possible Community Affiliation Matrices (c.f. Yang and Leskovec 2012).![Community detection method comparison for Link Clustering (L), Clique Percolation (C), Mixed-Membership Stochastic Block Model (M) and the Affiliation Graph Model (A) on different PINs (PPI) and other social networks. Methods are compared by the metric proposed by Ahn et al. [Yang & Leskovec 2012]](https://i0.wp.com/www.blopig.com/blog/wp-content/uploads/2014/10/Model_network_comparison_results.png?ssl=1)

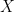 . In this article they proposed an exponential family of probability distributions to model
. In this article they proposed an exponential family of probability distributions to model 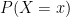 , where
, where  is a possible realisation of the random matrix
is a possible realisation of the random matrix  ) (see nodes 3-5 of the Figure below).
) (see nodes 3-5 of the Figure below).

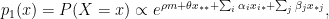
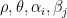 are parameters, and
are parameters, and  (identifying constrains).
(identifying constrains). 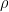 can be interpreted as the mean tendency of reciprocation,
can be interpreted as the mean tendency of reciprocation, 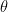 can be interpreted as the density (size) of the network,
can be interpreted as the density (size) of the network,  can be interpreted as as the productivity (out-degree) of a node,
can be interpreted as as the productivity (out-degree) of a node, 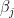 can be interpreted as the attractiveness (in-degree) of a node.
can be interpreted as the attractiveness (in-degree) of a node.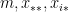 and
and 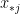 are: the number of reciprocated edges in the observed graph, the number of edges, the out-degree of node i and the in-degree of node j; respectively.
are: the number of reciprocated edges in the observed graph, the number of edges, the out-degree of node i and the in-degree of node j; respectively.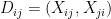 , the model assumes that all
, the model assumes that all  with
with 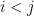 are independent.
are independent. and describe the joint distribution of
and describe the joint distribution of  , assuming all
, assuming all 
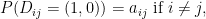
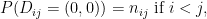
 .
.
 for
for 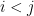 , and
, and 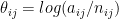 for
for 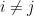 .
. and
and  can be interpreted as the reciprocity and differential attractiveness, respectively. With a bit of algebra we get:
can be interpreted as the reciprocity and differential attractiveness, respectively. With a bit of algebra we get:![exp(\rho_{ij})=[ P(X_{ij}=1|X_{ji}=1)/P(X_{ij}=1|X_{ji}=0) ]/[ P(X_{ij}=1|X_{ji}=0) / P(X_{ij}=0|X_{ji}=0) ],](https://s0.wp.com/latex.php?latex=exp%28%5Crho_%7Bij%7D%29%3D%5B+P%28X_%7Bij%7D%3D1%7CX_%7Bji%7D%3D1%29%2FP%28X_%7Bij%7D%3D1%7CX_%7Bji%7D%3D0%29+%5D%2F%5B+P%28X_%7Bij%7D%3D1%7CX_%7Bji%7D%3D0%29+%2F+P%28X_%7Bij%7D%3D0%7CX_%7Bji%7D%3D0%29+%5D%2C+&bg=ffffff&fg=000&s=0&c=20201002)

 , and
, and  where
where