
Last week I presented a paper by Klei et al. on a new module in the Phenix software suite. This module, entitled Guided Ligand-Replacement (GLR), aims to make it easier to place ligands during the crystallographic model-building process by using homologous models of the ligand-protein complex for the initial placement of the ligand.
In the situation where ligands are being added to a crystallographic protein model, a crystallographer must first build the protein model, identify the difference electron density, and then build the ligand into this density.
The GLR approach is particularly helpful in several cases:
- In the case of large complex ligands, which have many degrees of freedom, it can take a long time to fit the ligand into the electron density. There may be many different conformations of the ligand that fit the difference electron density to a reasonable degree, and it is the job of the crystallographer to explore these different conformations. They must then identify the true model, or perhaps an ensemble of models in the case where the ligand is mobile or present in different, distinct, binding modes. GLR makes this process easier by using a template from a similar, previously-solved structure. The ligand position and orientation is then transplanted to the new structure to give a starting point for the crystallographer, reducing the tedium in the initial placing the ligand.
- In the case of a series of related crystal structures, where the same protein structure is determined a number of times, bound to different (but similar) ligands. This is common in the case of structure based drug-design (SBDD), where a compound is developed and elaborated upon to improve binding affinity and specificity to a particular protein. This process generates a series of crystal structures of the protein, bound to a series of ligands, where the binding modes of the ligands are similar in all of the structures. Therefore, using the position and orientation of the ligand from a structure is a good starting point for the placement of further elaborations of that ligand in subsequent structures.
- In the case of several copies of the protein in the asymmetric unit cell of the crystal. After one copy of the ligand has been built, it can be quickly populated throughout the unit cell, removing the need for the crystallographer to undertake this menial and tedious task.
Program Description:
The required inputs for GLR are standard, as required by any ligand-fitting algorithm, namely:
- The APO structure of the protein (the structure of the protein without the ligand)
- A description of the ligand (whether as a SMILES string, or as a cif file etc)
- An mtz file containing the experimental diffraction data
Overview of the program:

Fig 1. Program Overview.
> Identification of the reference structure
Firstly, the program must determine the reference structure to be used as a template. This can be specified by the user, or GLR can search a variety of sources to find the best template. The template selection process is outlined below. Reference structures are filtered by the protein sequence identity, similarity of the molecular weights of the ligands, and finally by the similarity of the binary chemical fingerprints of the ligands (as calculated by the Tanimoto coefficient).

Fig 2. Reference Structure selection flow diagram.
Little justification is given for these cutoffs, although it is generally accepted that proteins with above 70% sequence identity are highly structurally similar. The Tanimoto coefficient cutoff of 0.7 presumably only serves to remove the possibly of very low scoring matches, as if multiple potential reference structures are available, the highest Tanimoto-scored ligand-match is used. They do not, however, say how they balance the choice in the final stage where they take the ligand with the highest Tanimoto score and resolution.
The method for assigning the binary chemical fingerprints can be found here (small error in link in paper).
> Superposition of Reference and Target structures
Once a reference structure has been selected, GLR uses graph-matching techniques from eLBOW to find the correspondences between atoms in the reference and target ligands. These atomic mappings are used to orient and map the target ligand onto the reference ligand.
Once the reference protein-ligand structure is superposed onto the target protein, these atomic mappings are used to place the target ligand.
The target complex then undergoes a real-space refinement to adjust the newly-placed ligand to the electron density. This allows the parts of the target ligand that differ from the reference ligand to adopt the correct orientation (as they will have been orientated arbitrarily by the graph-matching and superposition algorithms).
> Summary, Problems & Limitations
GLR allows the rapid placement of ligands when a homologous complex is available. This reduces the need for computationally intensive ligand-fitting programs, or for tedious manual building.
For complexes where a homologous complex is available, GLR will be able to quickly provide the crystallographer with a potential placement of the ligand. However, at the moment, GLR does not perform any checks on the validity of the placement. There is no culling of the placed ligands based on their agreement with the electron density, and the decision as to whether to accept the placement is left to the crystallographer.
As the authors recognise in the paper, there is the problem that GLR currently removes any overlapping ligands that are placed by the program. This means that GLR is unable to generate multiple conformations of the target ligand, as all but one will be removed (that which agrees best with the electron density). As such, the crystallographer will still need to check whether the proposed orientation of the ligand is the only conformation present, or whether they must build additional models of the ligand.
As it is, GLR seems to be a useful time-saving tool for crystallographic structure solution. Although it is possible to incorporate the tool into automated pipelines, I feel that it will be mainly used in manual model-building, due to the problems above that require regular checking by the crystallographer.
There are several additions that could be made to overcome the current limits of the program, as identified in the paper. These mainly centre around generating multiple conformations and validating the placed ligands. If implemented, GLR will become a highly useful module for the solution of protein-ligand complexes, especially as the number of structures with ligands in the PDB continues to grow.





 ) that maximises the following posterior distribution for the number of bins:
) that maximises the following posterior distribution for the number of bins:
 is the number of bins,
is the number of bins,  is the data,
is the data, 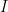 is prior knowledge about the problem, i.e. in particular the use of equal length bins and the range of data
is prior knowledge about the problem, i.e. in particular the use of equal length bins and the range of data 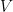 , which has the relation
, which has the relation  where
where  is the width of bins,
is the width of bins, 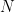 is the number of data points and
is the number of data points and 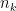 is the number of observations that fall in the
is the number of observations that fall in the 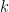 th
th  ) of the bins of the histogram is given by:
) of the bins of the histogram is given by: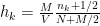 .
.




{kind=link}